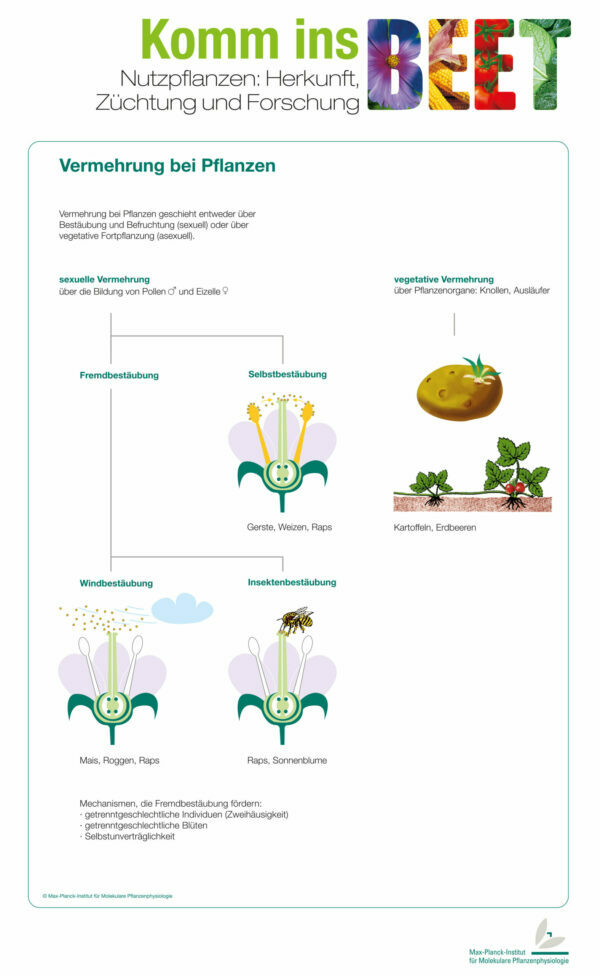
Blotting: Western, Norther, Southern Blot
Mit Hilfe der verschiedenen Blotting-Methoden ist es Möglich DNA (Southern Blot), RNA (Northern Blot) oder ganzen Proteinen (Western Blot) von einem Elektrophorese-Gel auf eine Zielmembran zu übertragen, um sie biochemisch nachzuweisen. Hiermit kann man diese Moleküle nun identifierzieren und quantifizieren. Die Gel-Elektrophorese erklären wir euch in diesem Beitrag: Gel-Elektrophorese.
Anhand der Forschungsarbeit unserer Doktorandin Mercedes wollen wir euch im folgenden die drei Methoden genau erklären und vorstellen.
Um die Fotosynthese bei Pflanzen besser zu verstehen, hat Mercedes – Doktorandin am Max-Planck-Institut für molekulare Pflanzenphysiologie in Potsdam-Golm – in eine Tabakpflanze ein Gen eingebracht, das die Fotosynthese verbessern könnte. Wie sie das Gen in die Pflanze einbringt und wie sie in ihrer Forschung vorgeht, hat sie in dem Film „Vom Gewächshaus ins Labor“ bereits gezeigt.
Nun muss sie schauen, ob das Gen, das sie interessiert tatsächlich in das Erbgut der Pflanze eingebracht wurde, ob es dort aktiv ist und ob es last but not least auch tatsächlich das entsprechende Protein bildet. Dazu trennt sie zunächst jeweils die verschiedenen Bestandteile (DNA, RNA, Proteine) durch Gel-Elektrophorese nach ihrer Größe auf. Die aufgetrennten Moleküle macht sie anschließend durch die Methode des Southern, Northern bzw. Western Blot sichtbar. So kann sie feststellen, ob das Gen ins der pflanzliche Genom integriert ist, ob es aktiv ist und ob auch tatsächlich das entsprechende Protein gebildet wird. Ist dies der Fall, so kann sie weiter untersuchen, welche Wirkung das eingebrachte Gen auf die Fotosynthese hat, ob es tatsächlich die Fotosynthese verbessert oder eher das Gegenteil der Fall ist.
Die Bezeichnungen Southern, Northern und Western Blot haben nichts mit den Himmelsrichtungen zu tun. Edwin Southern entwickelte in den 70er Jahren die molekularbiologische Methode zur Untersuchung von DNA-Sequenzen. In Anspielung an seinen Namen wurde der Nachweis von RNA-Sequenzen mit Northern Blot bezeichnet und der Nachweis von Proteinen als Western Blot.
In unserer Video-Reihe „Pimp your brain“ erklären Wissenschaftler des Max-Planck-Institut für Molekulare Pflanzenphysiologie, wie sie bei ihrer Forschung vorgehen.
Weitere Videos findet Ihr auf unserem Youtube-Kanal!
Weitere Infos zum Max-Planck-Institut für Molekulare Pflanzenphysiologie gibt es unter http://www.mpimp-golm.mpg.de.