Die heutigen Tomatenpflanzen mit ihren großen roten Früchten sind das Produkt langjähriger intensiver Zuchtarbeit. Ursprünglich stammt die Tomate aus Südamerika und wurde von den Spaniern nach Europa gebracht. Hier wurde über Generationen hinweg der leicht bittere Geschmack der Früchte ausgemerzt und die Tomate eroberte sich einen wichtigen Platz in unserem Speiseplan.
Bei der Sortenzüchtung durch Kreuzung und Auslese sind mit der Zeit viele wichtige Eigenschaften der Wildtomatenpflanze aus dem Genpool der Kultursorten eliminiert worden. So kommt es, dass unsere heutigen Tomatenpflanzen extrem pflegebedürftig sind. Sie müssen ausreichend mit Wasser und Nährstoffen versorgt werden, denn mit Trockenheit oder Nährstoffmangel kommen sie gar nicht gut zurecht. Außerdem sind viele Sorten anfällig gegenüber Pilzkrankheiten wie Mehltau oder Virusinfektionen.
Wildtomatenpflanzen haben ein viel größeres genetisches Reservoir. Sie sind sehr gut an ihre trockenen Standorte angepasst und kommen auch mit hohen Salzgehalten zurecht. Außerdem hat man einige Arten identifizieren können, die Resistenzen gegenüber Pilzkrankheiten aufweisen. Bisher weiß man jedoch nicht, welche Gene für die unterschiedlichen Eigenschaften der Tomaten verantwortlich sind.
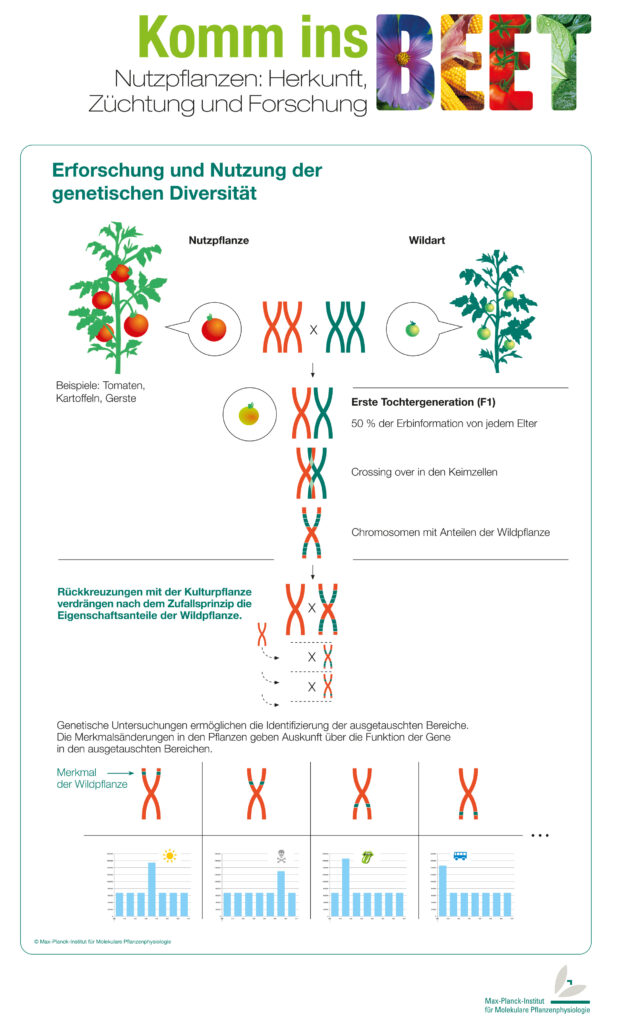
Introgressionslinien
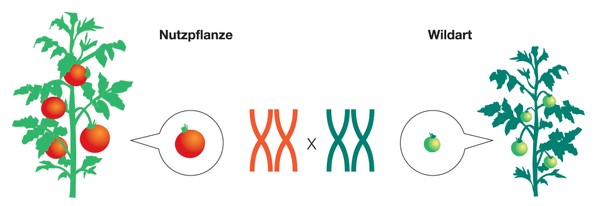
Bei der Erzeugung von Introgressionslinien werden einzelne Gene aus einer Art in eine verwandte Art überführt (lat. Introgression = Einkreuzung). Ausgangsmaterial für Introgressionslinien sind je eine Wild- und eine Kulturpflanzenart, die miteinander gekreuzt werden.
Die daraus resultierende Tochtergeneration hat ein Genom, das zu jeweils 50 Prozent aus Wild- und Kulturtomatenchromosomen besteht.
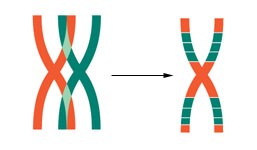
Während der Keimzellbildung der F1-Pflanzen kann es vorkommen, dass die Chromosomen Teilstücke untereinander austauschen (Crossing over). Dabei entstehen Chromosomen, die teilweise aus Kulturtomatengenen und teilweise aus Wildtomatengenen bestehen.
Die Pflanzen der F1-Generation werden jetzt mehrmals mit der Kulturtomatenpflanze gekreuzt. Dieser Vorgang heißt Rückkreuzung und dient dazu, den Anteil des Wildpflanzen-Genoms auf kurze Bereiche zu minimieren. Die dabei entstehenden Linien werden Introgressionslinien genannt und können teilweise neue, mitunter bessere, Eigenschaften aufweisen als die ursprüngliche Kulturpflanze.
Durch Sequenzanalyse kann man herausfinden, welche Abschnitte der DNA aus dem Genom der Wildpflanze stammen. Mit dieser Methode finden Forscher heraus, in welchen Bereichen der DNA bestimmte Eigenschaften verschlüsselt werden.
Bei der Erzeugung von Introgressionslinien werden einzelne Gene aus einer Art in eine verwandte Art überführt (lat. Introgression = Einkreuzung). Ausgangsmaterial für Introgressionslinien sind je eine Wild- und eine Kulturpflanzenart, die miteinander gekreuzt werden.
Viele Nutzpflanzen werden von Schädlingen angegriffen. Besonders der Mais hat unter dem Maiszünsler und dem Maiswurzelbohrer zu leiden. Da Insektenvernichtungsmittel nur begrenzt wirksam sind, haben Forscher der Maispflanze Selbstverteidigung beigebracht.
Unsere DNA ist natürlichen Mutationen unterworfen. Trotz sehr guter Reparaturmechanismen kann es zum Beispiel bei der Zellteilung vorkommen, dass eine falsche Base in die DNA eingebaut wird. Manche dieser Mutationen haben keinerlei Auswirkungen, sie werden „Stumme Mutationen“ genannt. Es kommt aber auch dazu, dass veränderte Proteine produziert werden, die der Zelle neue Eigenschaften verleihen, oder dass ein Gen so verändert wird, dass es kein funktionsfähiges Protein mehr produzieren kann. Diesen Effekt macht man sich bei der Mutationszüchtung zunutze.
Bei der Mutationszüchtung werden durch mutagene Chemikalien wie EMS (Ethylmethansulfonat) zufällig Punktmutationen ins Genom eingefügt. Da die DNA dabei unkontrolliert verändert wird, sind viele der so erzeugten Mutanten nicht lebensfähig oder weisen unerwünschte Veränderungen auf. Sie müssen dann aufgrund von Defekten entsorgt werden. Natürlich treten auch erwünschte Mutationen auf, die den Pflanzen neue, bessere Eigenschaften verleihen. Wenn solch eine Mutation identifiziert worden ist, muss sie durch Kreuzung wieder in eine leistungsfähige Zuchtlinie überführt werden.
Wie findet man die Mutationen?
Wenn sich eine falsche Base in der DNA befindet, kann der komplementäre Strang sich nicht mehr korrekt anlagern und die DNA bildet an dieser Stelle einen Buckel aus. Bestimmte Enzyme, wie zum Beispiel das Cel1, erkennen die Buckel und schneiden die DNA an den verformten Stellen durch. Es entstehen also mehrere kleine DNA-Abschnitte.
Mit Hilfe von Gelelektrophorese kann man die Länge der DNA-Stücke sichtbar machen. Sind alle DNA-Stränge gleich lang, so hat das Enzym keine Verformung erkannt und es liegt keine Mutation vor. Treten DNA-Stränge unterschiedlicher Länge auf, zeigt das eine Aktivität des Schneideenzyms an. Man ist auf eine Mutation gestoßen. Diese Methode wird TILLING genannt.
Mutationszüchtung unterliegt keinen gesetzlichen Regelungen, da keine Gene aus anderen Organismen eingeführt werden, sondern die natürliche Mutationsfrequenz erhöht wird. Auf diese Weise ist zum Beispiel eine Kartoffel gezüchtet worden, die keine Amylose mehr produziert. Das Gen für Amylose ist durch eine zufällige Mutation abgeschaltet worden.
Die Gel-Elektrophorese ist eine in der Molekularbiologie, Biochemie und Lebensmittelanalytik häufig eingesetzte Methode um verschiedene Moleküle voneinander zu trennen. Mercedes, Doktorandin am Max-Planck-Institut für molekulare Pflanzenphysiologie erklärt, wie die Gelelektrophorese funktioniert und wozu sie diese Methode für ihre Forschung einsetzt.
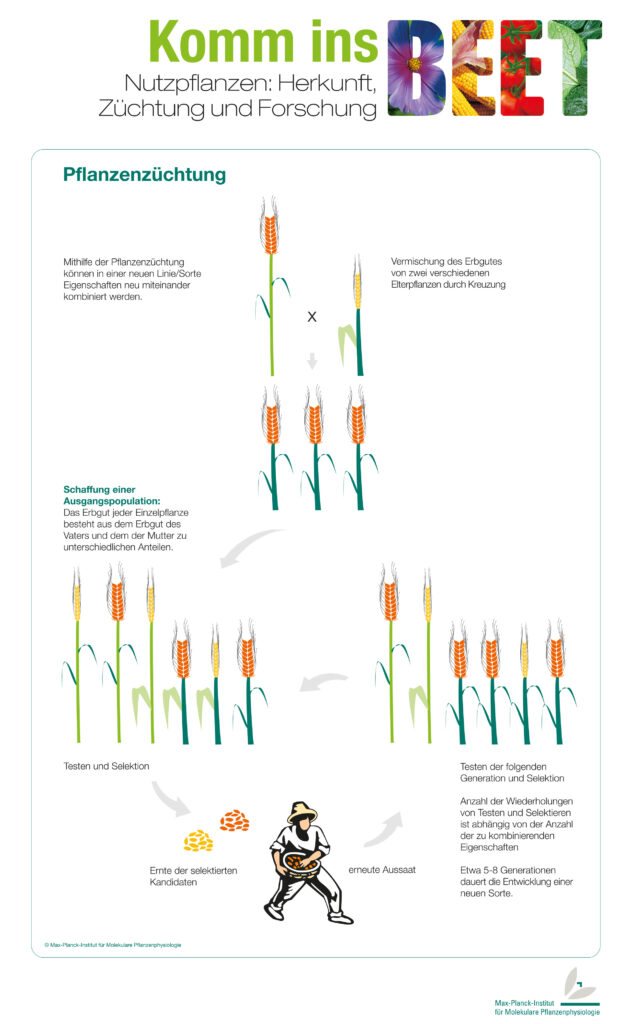
Die Auslese- und die Kombinationszüchtung sind die ältesten Formen der Pflanzenzüchtung. Dabei werden aus Wildpflanzen diejenigen mit vorteilhaften Eigenschaften ausgewählt und nur ihre Samen für eine erneut Aussaat verwendet. Durch das Auftreten von zufälligen Mutationen oder das Einkreuzen anderer Arten ist zum Beispiel aus dem Einkorn unser heutiger Weizen entstanden.
Zu Beginn der Steinzeit lebten Menschen als Jäger und Sammler. In der Epoche, die man heute als Jungsteinzeit bezeichnet, wurden sie sesshaft und es traten erste Formen von Ackerbau und Viehzucht auf.
Schon bald fiel den ersten Ackerbauern auf, dass manche Getreidepflanzen bessere Eigenschaften aufwiesen als andere. Im nächsten Jahr nutzten sie dann nur die Samen dieser Pflanzen zur Aussaat. Alle anderen wurden von der Vermehrung ausgeschlossen. Durch diese Auslese entstanden, über viele Generationen hinweg, so die ersten kultivierten Nutzpflanzen.
Aber nicht immer reicht eine reine Auslese der besten Vertreter einer Art aus um zum gewünschten Zuchtziel zu gelangen. Manchmal hilft erst eine Kombination aus Genen verschiedener Pflanzen. Das kann zufällig passieren oder vom Menschen herbeigeführt werden. Gemäß der ersten Mendel’schen Regel sind die Nachkommen der ersten Tochtergeneration (F1-Generation) uniform. Das bedeutet, dass sie alle die gleichen Merkmale besitzen. Problematisch wird es in der Enkel- und Urenkelgenerationen, hier spalten sich die Merkmale wieder auf. Deshalb ist es notwendig, über mehrere Generationen hinweg die geeigneten Nachkommen zu selektieren (auszuwählen).
Ziele bei der Züchtung von Nutzpflanzen
1. Vergrößerung des nutzbaren Pflanzenorgans (größere Körner, größere Samen)
2. stabile Ährenspindel bzw. keine automatische Öffnung der Schoten
3. Gleichzeitige Reifung aller Samen oder Früchte
4. Elimination der Samenruhe (Dormanz)
5. Aufrechtes Wachstum
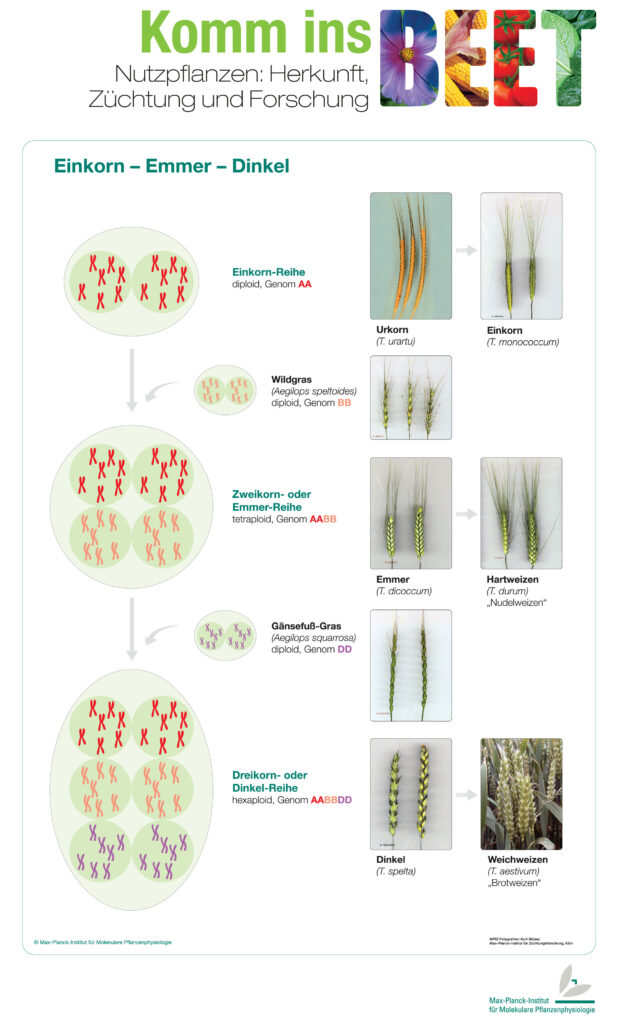
Vom Einkorn zum Weizen
Eine der ersten kultivierten Getreidearten war das Wildeinkorn (Triticum urartu). Es ist ein anspruchsloses Süßgras mit guter Krankheitsresistenz. Jedoch bildet Wildeinkorn nur wenige kleine Körner und hat eine brüchige Ährenspindel. Durch beständige Auslese der besten Einzelpflanzen züchteten die Menschen aus dem Wildeinkorn das kultivierte Einkorn (Tritium monococcum) mit größeren Körnern und fester Ähre.
Diese Art der Auslese allein hätte aber nicht ausgereicht um aus dem Einkorn unseren heutigen Weizen zu züchten. Dabei kam unseren Vorfahren auch der Zufall zu Hilfe.
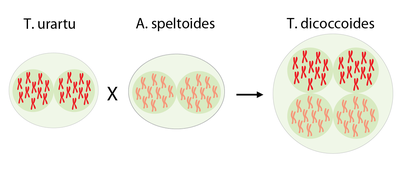
Durch spontane – also zufällige – Kreuzung mit einem anderen wilden Gras (Aegilops speltoides)* entstand der wilde Emmer (Triticum dicoccoides), auch Zweikorn genannt. Die beiden Elternlinien besaßen jeweils einen diploiden Chromosomensatz; jedes Chromosom lag doppelt im Zellkern vor. Bei der Kreuzung kam es zu einer Verdoppelung der elterlichen Chromosomensätze, sodass der Emmer tetraploid ist. Organismen, in denen Chromosomensätze aus unterschiedlichen Arten vorliegen, bezeichnet man als allopolyploid.
Aus dem wilden Emmer wählten Ackerbauern über viele Generationen hinweg wiederum die besten Pflanzen aus und es entstand der kultivierte Emmer (Triticum dicoccum). Dieser Emmer ist ein Vorläufer der ebenfalls tetraploiden Weizenart Hartweizen (Triticum durum), die besonders wichtig für die Herstellung von Nudeln und ähnlichen Teigwaren ist.
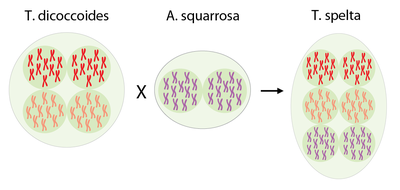
Wirtschaftlich und für unsere Ernährung bedeutsamer ist jedoch Weichweizen, der sogar einen hexaploiden Chromosomensatz aufweist. Der zusätzlich diploide Chromosomensatz kam vermutlich durch eine spontane Kreuzung mit dem diploiden Gras Aegilops tauschii* in das Genom. Dabei entstand zunächst der Dinkel (Triticum spelta) aus dem durch weitere Züchtung der Weichweizen (Triticum aestivum) entstand. Weichweizen ist ein wichtiger Ausgangsstoff für die Produktion von Brot und anderen Backwaren. Inzwischen ist Weizen nach Mais das am zweithäufigsten angebaute Getreide der Welt.
*wird manchmal als Triticum speltoides, manchmal als Aegilops speltoides bezeichnet, das gleiche bei Triticum/Aegilops squarrosa
Hier erklärt Eicj Kathleen, wie durch zufällige Einkreuzung von Wildgräsern unser Weizen entstanden ist.
Wir wissen, dass Weizen zum Brot backen verwendet wird oder aus Hartweizen Nudeln hergestellt werden, aber wer weiß schon, dass Weizen vor langer, langer Zeit aus Wildgräsern entstanden ist?
Was ist Triticale?
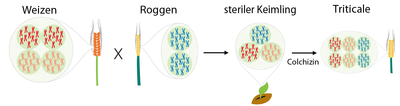
Die Getreideart Triticale ist vielen unbekannt. Dabei ist Triticale nichts weiter als eine Kreuzung aus Weizen und Roggen. Ziel dieser Kreuzung war es, die guten Eigenschaften beider Arten zu vereinen und die schlechten Eigenschaften auszulöschen.
Roggen hat keine hohen Ansprüche an Boden oder Klima und wächst auch auf nährstoffarmen Böden relativ gut. Dafür liefert er aber im Vergleich zum Weizen niedrigere Ernteerträge.
Weizen braucht einen ausgezeichneten Boden und viel gutes Wetter, bringt dafür aber auch viele qualitativ hochwertige Körner mit guten Klebeeiweißen (wichtig beim Backen) hervor.
Bestäubt man Weizenblüten mit dem Pollen von Roggenpflanzen so entsteht Triticale. Der Name dieser künstlichen Getreideart setzt sich aus den lateinischen Namen der beiden Elternpflanzen zusammen: Triticum, der Weizen, und Secale, der Roggen.
Jedes Elternteil vererbt die Hälfte seines Chromosomensatzes an die Nachkommen weiter. Das Problem hierbei ist, dass der zur Kreuzung verwendete Hartweizen tetraploid ist, also jedes Chromosom vier Mal besitzt, und der Roggen nur diploid, also mit je zwei Chromosomensätzen ausgestattet ist. Die Triticale erbt demzufolge drei Chromosomensätze, zwei vom Weizen und einen vom Roggen. Triploide Pflanzen wie Triticale sind oft unfruchtbar, weil bei einer Zellteilung die Chromosomen, aufgrund ihrer ungeraden Anzahl, nicht gleichmäßig auf die Keimzellen aufgeteilt werden können.
Deshalb unterzog man die Keimlinge einer Colchizinbehandlung. Dadurch wurde der Chromosomensatz in den Keimzellen verdoppelt und die Triticale besaß daraufhin einen sechsfachen Chromosomensatz. Damit ist sie fruchtbar und zur Ausbildung eigener Keimzellen fähig.
Vom Wildkohl zu den heutigen Kohlsorten
Rotkohl, Weißkohl, Blumenkohl, Brokkoli und Kohlrabi – dass diese Pflanzen verwandt sind, erkennt man inzwischen nur noch an ihren Namen. Ihr Aussehen ist komplett unterschiedlich und doch haben sie sich alle aus einer Wildart entwickelt: dem Gemüsekohl oder auch Wildkohl. Er stammt ursprünglich aus dem Mittelmeerraum und wächst vor allem an den Küsten Griechenlands, Italiens, Frankreichs und Spaniens.
Doch welche Pflanzenteile sind es eigentlich, die wir bei den einzelnen Kohlarten verzehren? Am einfachsten lässt sich diese Frage beim Grünkohl beantworten, von ihm verzehren wir die verdickten Blätter. Auch von Rotkohl, Weißkohl und Wirsing essen wir die Blätter und die Kohlrabiknollen sind nichts weiter als verdickte Sprossachsen. Doch um was handelt es sich bei Blumenkohl, Romanesco und Brokkoli? Hier landen die noch nicht voll entwickelten Blütenstände auf unseren Tellern. Wenn man die Pflanzen nicht rechtzeitig erntet, sondern weiter wachsen lässt, sieht man bald blühenden Kohl anstatt des typischen Kohlkopfes.
Der Chinakohl ist übrigens näher mit Raps und Rüben verwandt, als mit den übrigen Kohlsorten.
Alles über Kohl erzählt euch Kathleen und stellt Euch die verschiedenen Kohlsorten vor:
Weißkohl, Rosenkohl, Blumenkohl, Grünkohl oder Kohlrabi, bei all diesen Kohlsorten nutzen wir unterschiedliche Pflanzenteile für unsere Mahlzeiten. Welche Teile essen wir und wie sind die verschiedenen Kohlsorten überhaupt entstanden?
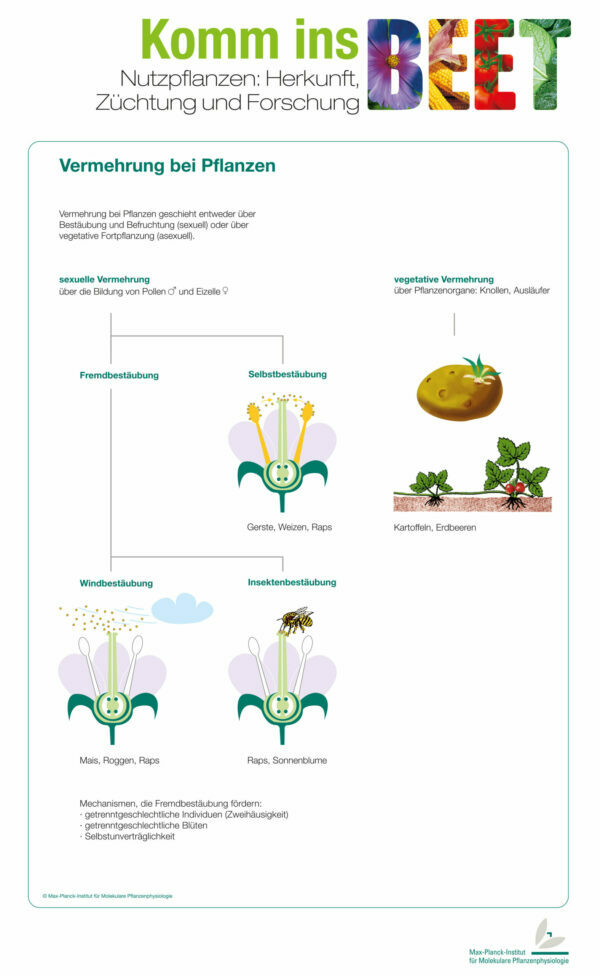
Einhäusig – zweihäusig – zwittrig? Pflanzen können verschiedene Blüten ausbilden und mit Hilfe von Wind oder Insekten ihren Pollen auf die Blütennarbe übertragen. Wer sich nicht auf Wind oder Bienen verlassen will, der kann wie die Erdbeere oder die Kartoffel auch auf vegetative Vermehrung zurückgreifen.
Die Vermehrung bei Pflanzen erfolgt entweder sexuell über Bestäubung und Befruchtung oder asexuell über vegetative Fortpflanzung. Bei der sexuellen Fortpflanzung werden in den Staubbeuteln der Pflanzen die männlichen Pollen und im Griffel die weiblichen Eizellen gebildet. Die Blüten verschiedener Arten sind dabei unterschiedlich zusammengesetzt.
Zwittrige Blüten vereinen weibliche und männliche Blütenorgane
Einhäusige Pflanzen haben getrennte weibliche und männliche Blüten auf derselben Pflanze (z.B. Mais)
Zweihäusige Pflanzen haben weibliche und männliche Blüten nur auf getrennten Pflanze (z.B. Brennnesseln)
Manche Pflanzen nutzen ihren eigenen Pollen zur Bestäubung der Narbe (z.B. Gerste und Weizen). Diese Selbstbestäubung hat den Vorteil, dass sich auch in verlassenen unwirtlichen Gebieten aus einer einzigen Pflanze eine ganze Population entwickeln kann.
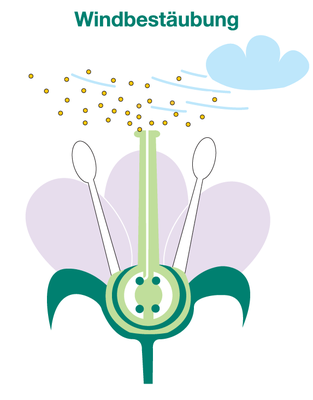
Andere Pflanzen sind Fremdbestäuber. Sie lassen ihren Pollen von Wind oder Insekten auf andere Pflanzen übertragen (z.B. Mais, Roggen und Sonnenblume). Fremdbestäubung hat den großen Vorteil, dass die Gene durchmischt werden und neue Kombinationen entstehen können, die den Nachkommen vielleicht bessere Eigenschaften verleihen.
Der Pollen von windbestäubten Pflanzen ist meist sehr leicht und klein, damit er sich weit verbreiten kann. Insektenbestäubte Pflanzen besitzen einen schwereren Pollen und zum Anlocken der Insekten besonders große und farbige Blüten.
Bei der vegetativen Vermehrung von Pflanzen werden auf verschiedenen Wegen Klone der Mutterpflanze gebildet, die zu selbständigen Pflanzen heranwachsen können. Dabei kann es sich um Ableger, Rhizome, Knollen oder andere Formen handeln. Ein Beispiel ist die Kartoffel, bei der aus den Augen der Knollen komplette neue Pflanzen heranwachsen können. Erdbeeren können lange Ausläufer bilden, an deren Enden neue Pflänzchen entstehen, die nach Absterben der Verbindung zur Mutterpflanze zu eigenständigen, genetisch identischen Pflanzen werden.
Dein Interesse ist geweckt und Du möchtest tiefer in das Thema einsteigen? Folgend nochmal alle Beiträge der Kategorie Züchtung:
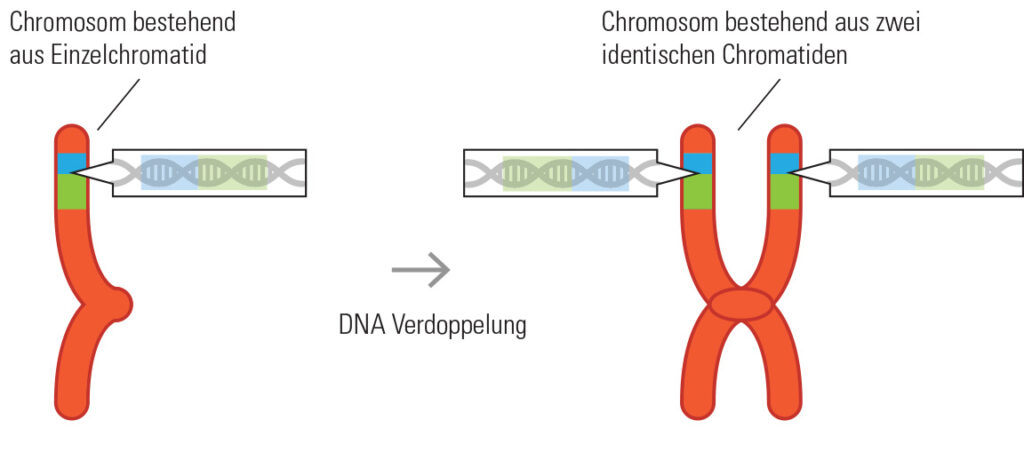
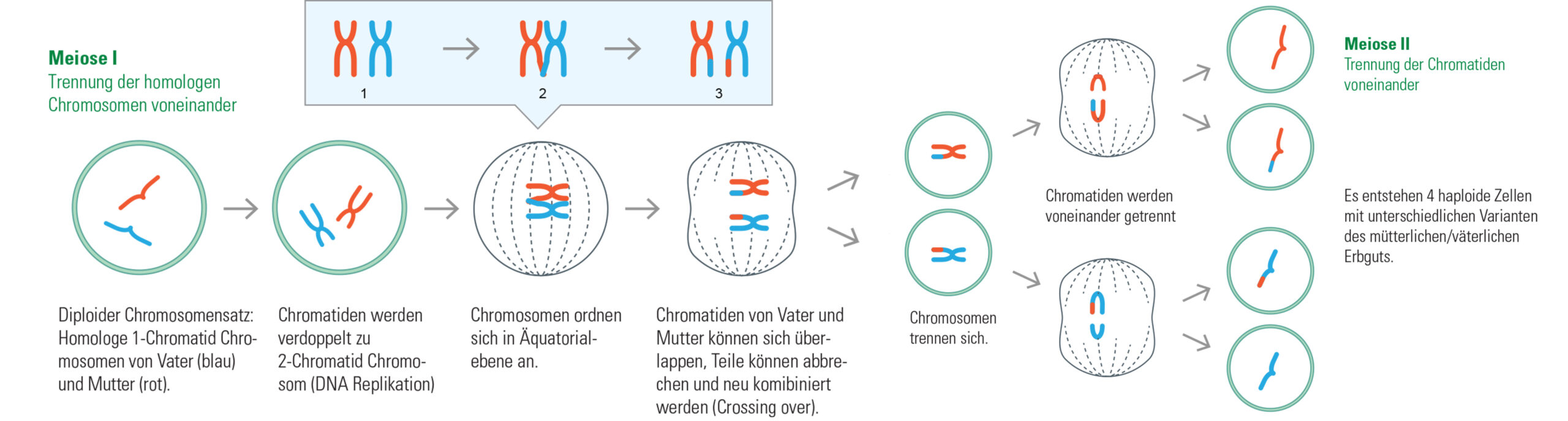
Wenn Zellen sich teilen, dann sollen beide Tochterzellen genau die gleiche Erbinformation enthalten, wie die Mutterzelle. Damit das klappt, müssen die Chromosomen zunächst einmal verdoppelt werden.
Wir haben gesehen, dass in einem Doppelstrang der DNA nur bestimmte Basen miteinander paaren können. Auch wenn nur ein Strang der DNA bekannt ist, ergibt sich daraus, wie der gegenüberliegende Strang aussehen muss. Genau diese Tatsache ist bei der Verdoppelung (Replikation) der DNA wichtig. Sie beginnt mit der Auftrennung des Doppelstrangs in zwei einzelne Stränge.
Jetzt können Enzyme genau die passenden Basen anlagern und somit zwei neue Doppelstränge erzeugen. Immer wenn die Enzyme Adenin erkennen, fügen sie auf der gegenüberliegenden Seite Thymin an. Bei Cytosin wird auf der anderen Seite Guanin eingebaut und so weiter. Die beiden DNA-Stränge, die auf diese Weise entstehen, werden Schwesterchromatiden genannt. Eigentlich sind sie sogar „Siaemesische Zwillingschromatiden“, denn sie gleichen sich wie ein Ei dem anderen und sind sogar am Zentromer miteinander verbunden. Die Phase des Zellzyklus, in der die DNA verdoppelt wird, wird Interphase genannt.
Abb.DNA-Verdoppelung Die meiste Zeit liegen im Zellkern fadenförmige 1-Chromatid Chromosomen vor. Die typischen x-förmigen Chromosomen sind nur sichtbar kurz vor bzw. während der Zellteilung (Mitose) bzw. vor der Reduktionsteilung (Meiose). Die übrige Zeit liegen die Chromosomen als diffus aufgelockerte 1-Chromatid-Chromosomen vor, wie dünne Fäden.
Die Mitose – aus eins mach zwei
Auf die Interphase folgt die Mitose, die eigentliche Kernteilung. Damit die langen DNA-Fäden dabei nicht beschädigt werden, werden sie zunächst kondensiert (verdichtet). Die Chromosomen sind dann unter dem Mikroskop als x-förmige Strukturen erkennbar. Den Mittelpunkt des X bildet das Zentromer. Als nächstes lagern sich die Chromosomen in der Äquatorialebene an, also in der Mitte des Zellkerns. Lange dünne Fäden, Spindelfasern genannt, greifen an den Zentromeren an und reißen die Schwesternchromatiden auseinander. Jeweils ein Chromatid jedes Chromosoms wird zu einem der Zellpole gezogen. Dort werden die Chromosomen wieder dekondensiert (entpackt) und es bildet sich eine neue Kernmembran aus. Anschließend teilt sich die Zelle.
Mitose
Die Meiose – eine besondere Art der Zellteilung
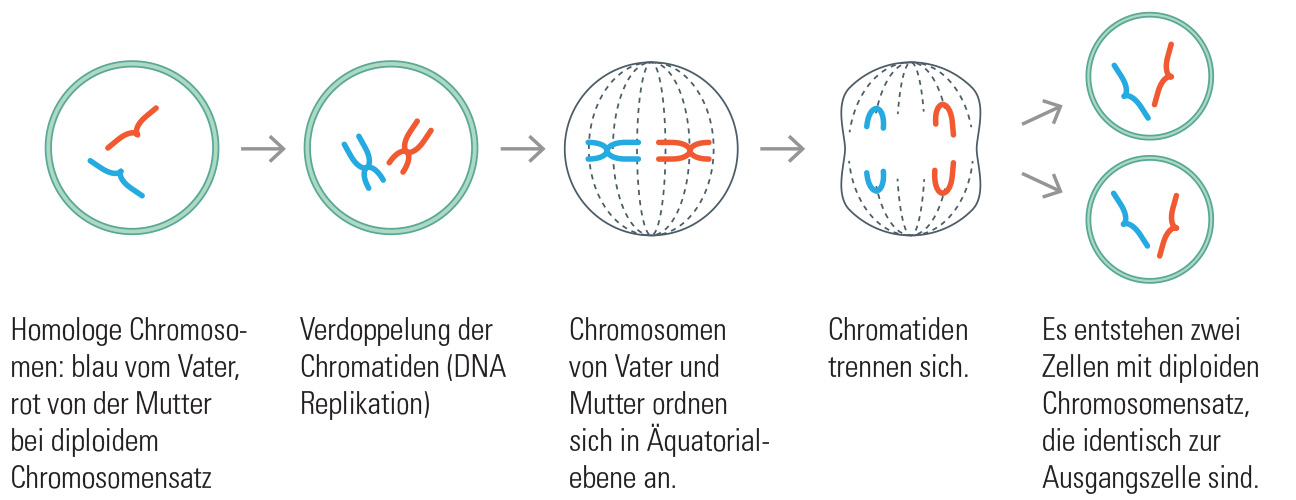
Bei geschlechtlicher Vermehrung geben die Eltern Erbinformation (Chromosomen) an ihre Kinder weiter. Damit sich nicht mit jeder neuen Generation die Anzahl der Chromosomen verdoppelt, werden Keimzellen gebildet, die nur über den halben Chromosomensatz verfügen. Die meisten Organismen sind diploid und ihre Keimzellen demzufolge haploid. Der Vorgang der Keimzellbildung wird Meiose genannt.
Bei der Meiose ordnen sich die kondensierten Chromosomen paarweise in der Äquatorialebene an. Jeweils ein väterliches und ein mütterliches Chromosom liegen nebeneinander. Genau wie bei der Mitose greifen Spindelfasern an den Zentromeren an. Statt einzelner Chromatide, werden ganze Chromosomen zu den beiden Zellpolen gezogen. Anschließend findet die Zellteilung statt, wie sie oben beschrieben ist.
Erst nach der ersten Teilung werden die Chromosom in ihre beiden Schwesterchromatiden zerlegt und es folgt eine erneute Zellteilung. Aus einer diploiden Zelle sind vier haploide Keimzellen entstanden, deren Erbinformation sich zufällig aus mütterlichen und väterlichen Anteilen zusammensetzt.
Im Dezember 2014 hat Max Heeke von detektor.fm das Max-Planck-Institut für Molekulare Pflanzenphysiologie besucht und sich mit unserem Direktor Prof. Mark Stitt über die Photosynthese unterhalten. Dabei ging es vor allem um die sogenannten C3- und C4- Pflanzen und wie deren Erforschung mit der Ernährung der Weltbevölkerung zusammenhängt.
Forschung für die Ohren
Den Podcast „Photosynthese gegen den Welthunger“ findet Ihr bei detektor.fm
In Koopertion zwischen Max-Planck-Gesellschaft und dem Online-Radio detektor.fm entstehen Forschungs-Podcasts zu den unterschiedlichsten Themen, die Ihr entweder über die Streamingdienste Deezer oder Spotify anhören könnt oder aber Ihr ladet Euch die Max-Planck-App für das iPhone im iTunes-Store herunter. Über die App erhaltet Ihr u.a. Zugriff auf die aktuellen Pressemeldungen der verschiedenen Max-Planck-Institute und deren Forschungsergebnisse.
Die meisten Pflanzen betreiben eine so genannte C3-Fotosynthese. Sie ist die älteste und bei gemäßigten Temperatur- und Lichtverhältnissen auch wirkungsvollere Art der Bindung des Kohlenstoffdioxids, das über die Spaltöffnungen passiv aufgenommen wird.
Wie in den Grundlagen erläutert, ist das Kernstück der lichtunabhängigen Reaktion, auch als Dunkelreaktion bezeichnet, das Enzym RuBisCo, das die Kohlenstoffdioxidfixierung katalysiert.
Es entstehen im ersten Schritt der CO2 -Fixierung im Calvin-Benson-Zyklus mit Hilfe des Enzyms RuBisCo aus einem Molekül Kohlendioxid und Ribulose 1,5-bisphosphat zwei Moleküle 3-Phosphoglycerat (3-PGA). Von dieser Verbindung, die aus drei Kohlenstoffatomen aufgebaut ist, leitet sich der Name C3-Fotosynthese ab und alle Pflanzen, die diese Art der Fotosynthese betreiben werden als C3-Pflanzen bezeichnet. 90 Prozent aller Landpflanzen gehören zu den C3-Pflanzen, wie z.B. Weizen und Reis. Erdgeschichtlich sind diese Pflanzen zuerst entstanden, da die Atmosphäre reich an CO2 war.
Was die Pflanzen nicht vorhergesehen hatten – Fehler im System
Vor ca. 4,6 Milliarden Jahren war in der Erdatmosphäre so gut wie kein Sauerstoff enthalten, dafür war die Kohlenstoffdioxidkonzentration hoch. Man vermutet bis zu ca. 20 Prozent. Durch die Entwicklung von fotosynthetisch aktiven Bakterien, Algen und später auch Pflanzen reicherte sich die Atmosphäre mit Sauerstoff an und Kohlenstoffdioxid nahm ab. Der Sauerstoffgehalt der Atmosphäre beträgt heute 21 Prozent, der CO2-Anteil liegt bei 0,04 Prozent. Diese Umkehrung der Verhältnisse blieb für die Fotosynthese nicht ohne Folgen.
Das Enzym RuBisCo, das für die CO2-Fixierung zuständig ist, kann nämlich sowohl mit CO2 als auch mit O2 reagieren. Durch die Abnahme des Kohlenstoffdioxids und die Sauerstoffanreicherung in der Atmosphäre, die die Pflanzen selber mit verursacht haben, sinkt die Effektivität der Fotosynthese, da die Wahrscheinlichkeit, dass das Enzym RuBisCo auf ein Sauerstoffatom stößt viel größer ist, als auf ein CO2-Atom zu stoßen.
Verstärkt wird dieses Problem bei Trockenheit bzw. steigenden Temperaturen. Die Spaltöffnungen des Blattes werden dann geschlossen, um Wasserverluste zu verringern. Deshalb gelangt weniger CO2 in die Zellen. Diese Situation führt insgesamt dazu, dass weniger Zucker entsteht, also die Fotosynthese weniger effektiv arbeitet.
Die Evolution „rettet“ ihre Kinder – Entstehung von C4– und CAM-Pflanzen
Bei den Pflanzen, die einen C3-Stoffwechsel betreiben, gelangt das Kohlenstoffdioxid passiv durch die Spaltöffnungen in die Zellen, um dann im Zuge des Calvin-Benson-Zyklus fixiert zu werden. Man nimmt heute an, dass der vor ca. 30 Millionen Jahren, während des Oligozäns, einsetzende Kohlenstoffdioxidrückgang die Grundvoraussetzung war für die Entwicklung bestimmter Pflanzen, deren Fotosynthese anders verläuft, als die der C3 Pflanzen. Nach einem ersten Evolutionsschub haben weiter sinkende CO2 Konzentrationen bei gleichzeitigem Anstieg der Sauerstoffkonzentrationen in der Atmosphäre und lokalen Faktoren, wie hohe Temperaturen, geringe Niederschlagsmengen und Trockenheit, sowie hoher Lichteinfall bei der Entstehung dieser Art von Pflanzen eine Rolle gespielt. Pflanzen des C4 Typs sind in 19 Pflanzenfamilien bis zu 61 Mal unabhängig voneinander entstanden. Es gibt sechs globale Zentren, die als Keimzelle für diese Pflanzen gelten: Nord- und Südamerika, Südafrika, Nordostafrika und Arabien, sowie Zentralasien und Australien.
Während in C3 Pflanzen die CO2-Fixierung und der sich daran anschließende Stoffwechsel, der zur Bildung von Zuckern führt in ein und demselben Zelltyp abläuft, sind bei C4 – bzw. CAM-Pflanzen diese Abläufe entweder räumlich oder zeitlich voneinander getrennt.
Einerseits nehmen C4 Pflanzen im Unterschied zu den C3 Pflanzen CO2 aktiv unter Energieverbrauch auf, das dann in den sogenannten Mesophyllzellen fixiert wird. Hierbei entsteht statt einer Verbindung, die aus 3 C-Atomen besteht (Phosphoglycerinsäure), eine Verbindung, die über 4 C-Atome verfügt (Oxalacetat). Das in Form der 4-C Verbindung gebundene CO2 wird über besondere Verbindungen in die angrenzenden Bündelscheidenzellen transportiert und wird dort freigesetzt. Es sammelt sich in diesen Zellen an, so dass hohe CO2 Konzentrationen entstehen, die ca. 10-mal höher sind als in der Außenluft und die so für die RuBisCo zur Verfügung stehen, die dort wie bei den C3 Pflanzen für die weitere Umsetzung sorgt aber mit starker Unterdrückung der verschwenderischen Nebenreaktion mit Sauerstoff. Durch den Trick der aktiven Aufnahme und der räumlichen Trennung der Stoffwechselwege ist die Fotosynthese dieser Pflanzen effizienter als in C3 Pflanzen, vor allem bei höheren Temperaturen. Das Temperaturoptimum dieser Pflanzen liegt bei 30 bis 47 Grad Celsius, während es bei C3 Pflanzen bei 15 bis 25 Grad Celsius liegt.
Bei zunehmender Trockenheit und höheren Temperaturen sind diese C4 Pflanzen unseren C3 Pflanzen überlegen. Die Pflanzen erreichen hohe Biomasseerträge. Zu den C4 Pflanzen gehören u.a. Mais, Zuckerrohr, Amarant und Hirse.
Aufnahme von Kohlenstoffdioxid während der Nacht – CAM-Pflanzen
Bei anderen Pflanzen, wie beispielsweise Kakteen geht es nicht um die räumliche Trennung dieser Abläufe, sondern um die zeitliche Trennung. So öffnen Wüstenpflanzen nachts ihre Spaltöffnungen, um Kohlenstoffdioxid aufzunehmen. Auch hier entsteht in einem ersten Schritt zunächst eine Verbindung mit 4 C-Atomen (Äpfelsäure). Diese wird nach einem weiteren Umbau in den Vakuolen der Zelle gespeichert. Tagsüber, wenn die Spaltöffnungen geschlossen sind um Wasser zu sparen, wird CO2 aus der in der Vakuole gespeicherten Äpfelsäure freigesetzt und kann dann für die Fotosynthese genutzt werden. Man nennt diese Pflanzen, zu denen beispielsweise Kakteen und Ananasgewächse gehören auch CAM-Pflanzen, wobei CAM für Crassulaceen-Säurestoffwechsel (engl. Crassulacean acid metabolism) steht.
Wenn Ihr mehr über die Fotosynthese lernen möchtet, schaut doch mal unter „pimp your brain“ Filme zur Fotosynthese auf unserem YouTube Kanal.
Prof. em. Dr. Mark Stitt erklärt die CAM-Fotosynthese, die z.B. von Kakteen genutzt wird. Unter extremen Umweltbedingungen – wie in der Wüste – müssen sich Pflanzen eines Tricks bedienen, um Fotosynthese betreiben und somit wachsen zu können, was sie im wahrsten Sinne des Wortes ziemlich sauer macht.
Pflanzen: sie sind grün und kommen in den verschiedensten Formen vor. Sie bilden die unterschiedlichsten Lebensräume und liefern so die Voraussetzung für die Artenvielfalt. Sie beeinflussen aufgrund ihrer Fotosynthese die Luftzusammensetzung und haben zusammen mit Bakterien menschliches und tierisches
Unsere Kulturpflanzen werden von einer ganzen Reihe Schädlingen heimgesucht, die die Ernten bedrohen und Hungersnöte auslösen können. Wann und wie hat man überhaupt herausgefunden, dass es Pilze, Bakterien und Viren gibt und welcher Zusammenhang zwischen ihnen und den bei Pflanzen, aber auch bei Menschen und Tieren auftretenden Krankheiten besteht?