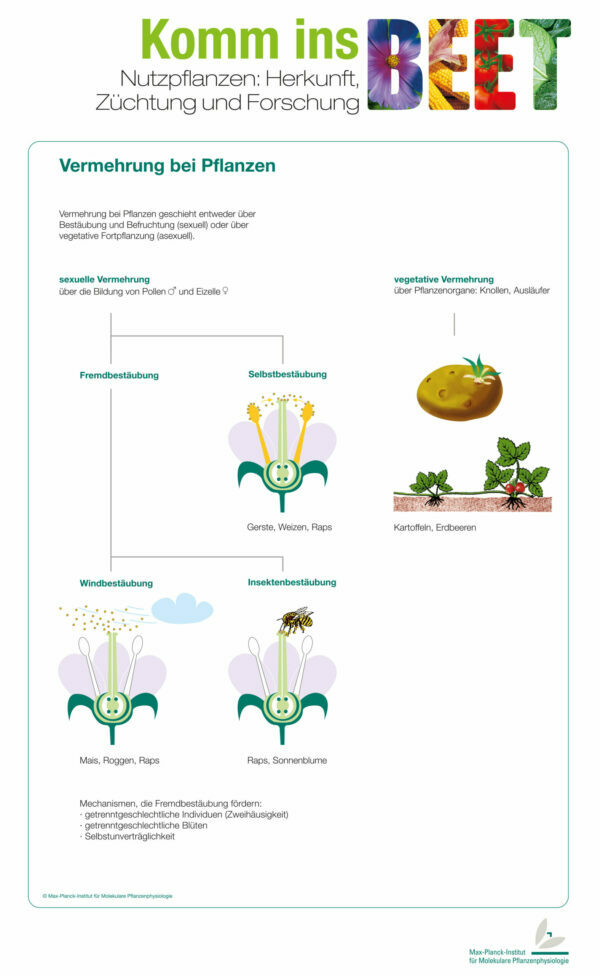
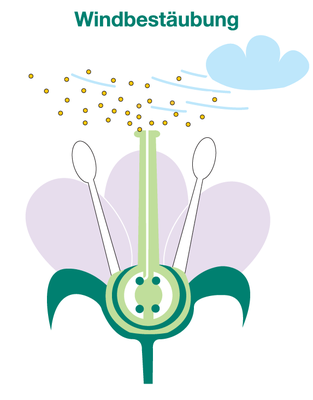
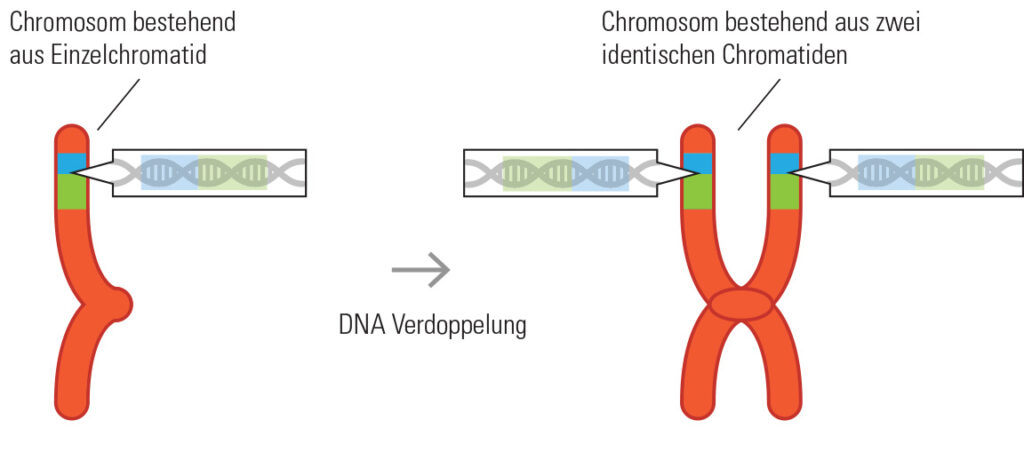
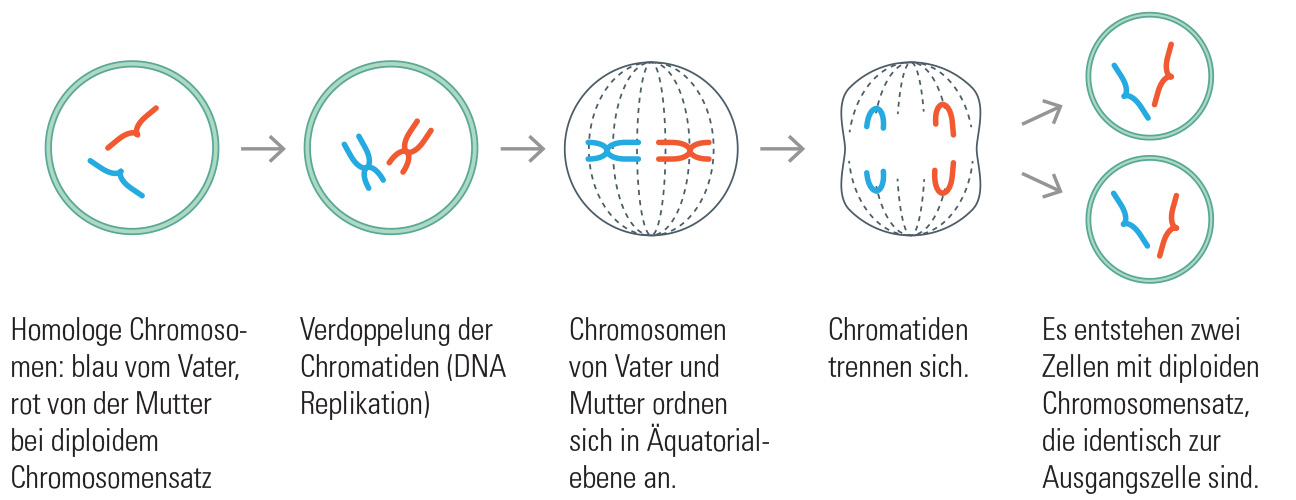
Biogas
Methan ist ein klimaschädigendes Treibhausgas. Verbrennt man jedoch Methan, wird dabei viel Energie frei. Diese Energie kann zur Erzeugung von Strom oder zur Erwärmung von Wasser genutzt werden.
Vom Mais zum Methan
In der Landwirtschaft fällt viel Abfall an: Klärschlamm, Pflanzenreste oder Kuhmist zum Beispiel. Bisher haben die Bauern diesen Müll zur Düngung ihrer Felder verwendet oder aufwendig entsorgt. In Biogasanlagen aber kann der Abfall zur Energieerzeugung verwendet werden. Bakterien vergären praktisch alle Biomasse zu Gas, das zum größten Teil aus Methan besteht. Methan ist ein energiehaltiger Kohlenwasserstoff, bei seiner Verbrennung entsteht viel heiße Luft, die über eine Turbine Strom erzeugen und außerdem auch Wasser erwärmen kann. Die entstehende Wärme wird meist direkt vor Ort verbraucht, also in den Haushalten, die dem Biomasseheizkraftwerk (BMHK) am nächsten sind. Der Strom jedoch wird in das Stromnetz eingespeist und steht jedem Endverbraucher zur Verfügung.
Der große Vorteil von Biogas ist, dass praktisch alle zur Erzeugung notwendigen Rohstoffe Abfall sind und daher kostenlos anfallen. Außerdem stehen Bioabfälle das ganze Jahr über zur Verfügung und die Stromerzeugung aus Biogas ist daher – anders als bei Solarzellen und Windrädern – nicht jahreszeitenabhängig. Kritisiert wird bisher vor allem, dass auch solche Biomasse verbrannt wird, die mit schädlichen organischen Stoffen behaftet ist, wie zum Beispiel lackiertes Holz. Daher versuchen die Forscher gezielt Energiepflanzen zu entwickeln, die bei der Verbrennung eine hohe Energieausbeute erzielen, z.B. Energiemais.


Energiemais
Maisfelder dominieren in vielen Regionen inzwischen die Felder, obwohl der Verbrauch an Mais als Futter- oder Nahrungsmittel nicht gestiegen ist. Wer also hat so großen Hunger auf Mais? Es sind vor allem die Biogasanlagen, in denen die Maispflanzen von Bakterien zu Methan vergoren und so zur Energieerzeugung verwendet werden. Im Vergleich zu den anderen Getreidearten wie Weizen, Roggern oder Gerste liefert Mais tatsächlich die meiste Biomasse. Trotzdem wurde Mais jahrtausendelang als Nahrungs- und nicht als Energiepflanze kultiviert. Unsere Maispflanzen sind so gezüchtet, dass sie möglichst große Maiskolben mit vielen stärkehaltigen Körner hervorbringen. Das Zuchtziel, was bei Energiepflanzen jedoch an erster Stelle steht heißt: Biomasse. Der herkömmliche Mais soll also züchterisch so verändert werden, dass er weniger Kolben und mehr Blattwerk produziert.
Für eine Pflanze ist es energetisch sehr aufwändig, Samen zu produzieren. Körnermais ist züchterisch auf die Produktion von Samen, also Maiskörnern, optimiert. Energiemais soll jedoch keine Samen, sondern vor allem viel Pflanzenmasse liefern. Man muss den Energiemais also von der Blüte abhalten. Doch wie weiß eigentlich der Mais, wann die optimale Blütezeit gekommen ist?
Pflanzen haben eine innere Uhr. Anhand der Tageslänge wissen sie immer, welche Jahreszeit gerade herrscht und können ihre Stoffwechselvorgänge danach ausrichten. Das wird als Photoperiodismus bezeichnet. Dabei sind die Pflanzen sehr gut an ihre jeweiligen Standorte angepasst. Die Tage in Mitteleuropa sind länger als in südlichen Breitengraden, dafür haben wir weniger warme Sommermonate. Damit die Maispflanzen die warme Periode optimal zum Wachsen ausnutzen können, blühen sie in unseren Breitengeraden im Frühsommer. In Südeuropa sind die Tage im Sommer zwar etwas kürzer, dafür ist es bis in den Spätherbst hinein sehr warm. Die südeuropäischen Sorten blühen deshalb erst später im Jahr. Fügt man das Blüh-Gen aus den südlichen Sorten in die nördlichen ein, kommt die innere Uhr des Mais durcheinander. Dadurch kommt der Mais in unseren gemäßigten Breitengraden erst sehr spät oder gar nicht zur Blüte und verschwendet keine Energie auf die Produktion von Maiskolben. Stattdessen, so das ehrgeizige Ziel der Züchter, soll die Pflanze weiterwachsen, zwischen drei bis vier Meter hoch werden und somit etwa doppelt so viel Biomasse erzeugen wie normale Maissorten.